Rheotaxis in Zebrafish (Danio rerio)
While navigating a fluid environment like the water(swimming) or air (flying), an animal must learn to swim against the flow, and hold itself steady against the displacements induced by the surrounding air or water flow. From the animals frame of reference, these displacements can be detected as changes in their field of vision. However, even in the absence of such illuminating visual cues, aquatic animals seem to know how to orient themselves and swim against the oncoming flows. What navigational cues do they use, and how do they process it?
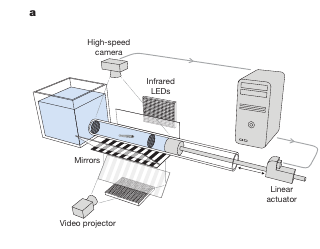
The first challenge one faces while trying to answer such questions is to replicate the niche of the model she is interested in. For example, to optimally extract rheotaxis behaviour from your average zebrafish, one needs to induce controlled laminar flow that would induce such behaviour in larvae.
Why use zebrafish?
Zebrafish live in the shallow waters of flood plains, paddies and streams in South Asia. Even in these gentle environments, the larva must hold its ground (or water :D) so not be swept away from its favourite microhabitat, to intercept drifting prey, to avoid predators by hiding in the dense vegetation, and navigate through its surroundings.
Interestingly, several species of predatory fishes and invertebrates prey on zebrafish using suction feeding, and the larva must be able to swim against this strong suction force if it has to escape.
While we know that the lateral line, the span of neuromasts on the fish body, and other such factors are important in mechanosensation, the sensory basis of navigation and the state dependent modulation of such behaviours are mostly unknown.
To investigate, Oteiza et al., built a specialized transparent tube to induce controlled laminar flows and used Particle Image Velocimetry (PIV) to map the flow gradients. They also mounted the entire setup on rails to physically accelerate the water and fish together, ruling out the possibility that the fish were simply detecting linear acceleration. In such situations it is important to remember that if a fish is being dragged by a homogenous current, it will be impossible to detect the oncoming waterflow because the fish will be navigating from withing the frame of reference of the moving water, and the dynamics will remain invariant under a velocity transformation.
Contact with the wall and external cues detected by organs like the inner ear might provide information from an extrenal frame of reference.
What physical cues do fish use to perform rheotaxis in the absence of visual information?
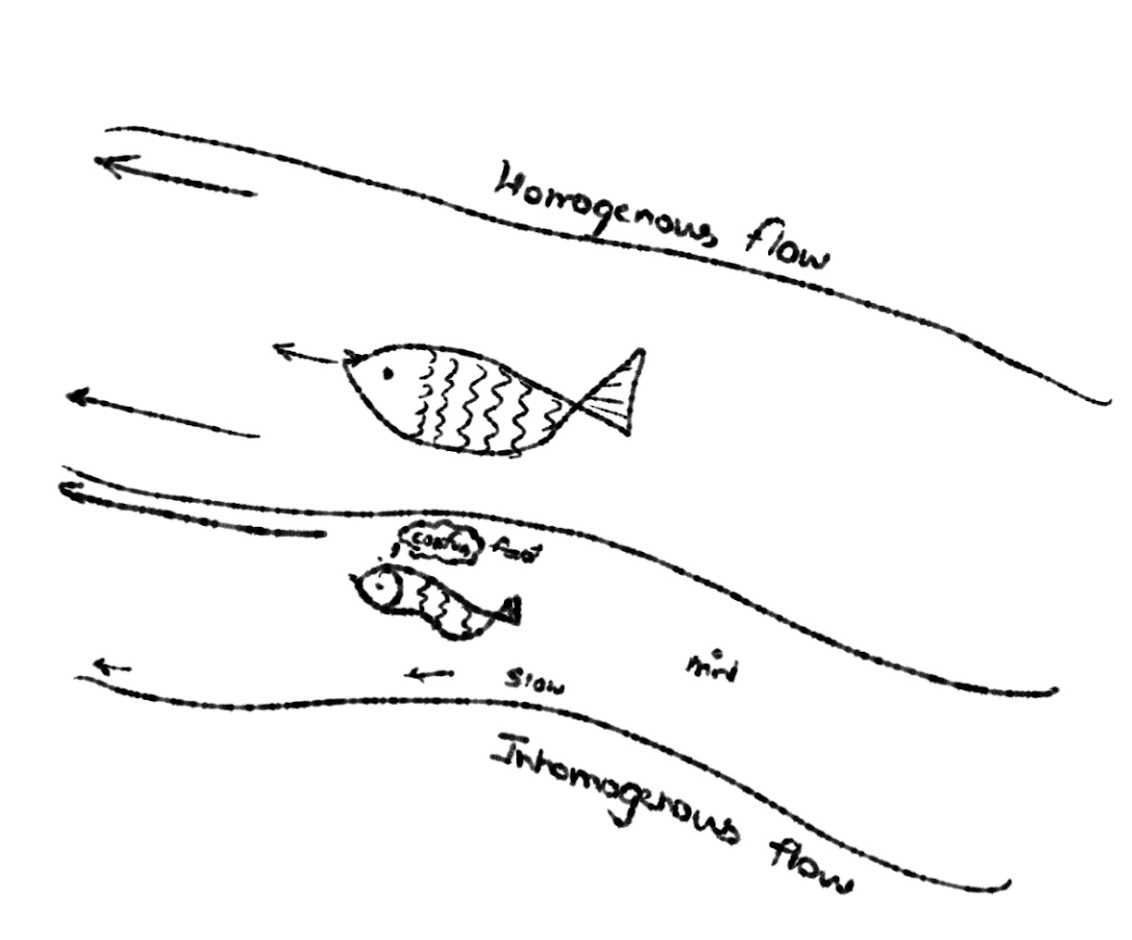
A perfectly uniform flow provides no directional cues to a drifitng larva. However, it has been demonstrated that fish bypass this by relying on the flow velocity gradient instead of the flow itself. The swimming fish, when subjected to a flow gradient in its environment, experiences a shear force (or curl) around its perimeter. if the fish was paralysed, this flow would succed in making the fish spin about its axis. Researchers proved that the ability to orient against a current improves with steeper gradients and disappears completely in zero-gradient (perfectly uniform) flows.
Mathematically, the rotational flow field is directly related to the local curl ($\nabla \times \mathbf{V}$) ff the water volume around the body by Stokes’ theorem:
$$ \iint_S (\nabla \times \mathbf{V}) \cdot d\mathbf{A} =\oint_{C} \mathbf{V} \cdot d\mathbf{s} $$
Thus, the fish can estimate the local curl in the water at its current position by calculating a lie integral of the relative water flow around its circuference.
Which specific sensory organs detect these flow gradients?
Several studies have confirmed the mechanosensory lateral line to be the primary sensor for rheotaxis, especially in the absense of visual cues.
The lateral line is the critical sensory network for vision-independent rheotaxis. It detects water flow relative to the body using arrays of receptor units called neuromasts, which consist of hair cells and support cells. This regions is responsible for sensing the flow gradient at a specific point.
The lateral line is functionally divided into anterior (head) and posterior (trunk and tail) regions. Studies manipulating these regions show that the posterior lateral line is fundamentally required for detecting continuous suction and orienting against flows. Caudal (tail) neuromasts appear especially important, likely because they connect directly to fast-swimming spinal motor networks that orchestrate escape bursts.
To accurately estimate the rotational flow fields necessary for gradient-based rheotaxis, fish must integrate sensory input from both sides of their body simultaneously. Unilateral disablement of the lateral line impairs rheotactic orientation just as severely as bilateral disablement.
Research indicates that the superficial neuromasts (located on the skin surface), rather than the canal neuromasts (enclosed in subcutaneous channels), specifically mediate this orienting behaviour.
Outside of early flow recognition and swimming in dark environments, the visual system (importantly, the optomotor reflex) plays an important role.
When both visual and lateral line cues are available, there is a clear prioritization rather than a weighted integration. The lateral line initiates the response, allowing the fish to trigger a counter-flow sequence earlier (further away from a suction source). But once counterflow swimming begins, the visual system completely overshadows the lateral line.
During visual mediation, the visual system dictates the precise swimming patterns, overriding the lateral line to tune the fish’s tail curvature and the number of tail beats needed to efficiently hold its position at varying distances from a flow source.
What are the behavioural mechanisms of rheotaxis?
Rheotaxis generally encompasses two main behavioral phases: initial orientation to the current and counterflow swimming to hold position. When first encountering an environment, larval zebrafish engage in an exploration phase, actively randomizing their body orientation. Once they detect the necessary flow cues, there is a clear and virtually instantaneous transition to counterflow swimming sequences (CSS), where they actively lock their orientation onto the local fluid streamlines.
To orient themselves and avoid being swept away, fish rely on a surprisingly simple behavioral algorithm based on temporal changes in water flow. Because larval zebrafish swim in discrete bursts (swim bouts) separated by resting periods, they constantly experience changes in velocity gradients. This change ($\Delta gradient$) defines the conditions in which the next bout will occur:
- when a fish experiences a decrease in the gradient magnitude, it subsequently swims straight (less flow-induced displacement)
- when a fish experiences an increase in gradient magnitude, it exhibits a high-magnitude turn towards the diection of the rotational field.
Physiological relevance:
Because the strongest velocity gradients naturally form near boundaries and obstacles, this >“gradient avoidance” behavior naturally drives the fish away from shorelines and toward the >center of the stream, where the flow velocity is highest but the gradient is flattest.
Such small changes in gradient magnitude mainly occur in the left-right axis of the body of the fish, and is independent of the flow direction and the passive rotations of the animals body. Additionally these changes allow the fish to orient both against and along the water flow.
What strategies do fish use to hold their position in a dynamic environment?
Once a fish begins counterflow swimming, the specific behavioral mechanism it uses to hold its position depends on which sensory systems are available:
- Lateral line mediation: When relying solely on hydromechanical cues (the lateral line), fish hold their position by stabilizing around a specific “operating point.” They exhibit repulsion when they get too close to the suction source (direction of the gradient) and attraction when they drift too far away. They tune this position by adjusting their average number of tail beats per bout and the resting periods between bout.
- Visual cues: When visual cues are available, zebrafish can efficiently hold their position at any distance from the source, provided they are beyond a minimum repulsion threshold. They adjust this by altering their tail curvature and the number of tail beats.
Escape and Avoidance Bursts
In the presence of dangerous, localized currents, such as a continuous point-source of suction, larval zebrafish execute a specific avoidance behavior. As they approach the suction, they initiate a sharp swimming burst to escape, often moving at least one-third of their body length away. The magnitude of this behavior is dynamically scaled: the closer the fish is to the suction source (where the flow is faster), the faster and farther it will burst. This could be a preprogrammed motor response scaled to the initial flow strength, or the result of continuous sensory feedback where the fish keeps bursting until it senses the flow has dropped to a safe level.
Researchers have also answered how behavioral compensation changes during early development (from 5 to 9 days post-fertilization). While the net result is always successful position holding, the mechanisms shift with age. In lateral-line-only conditions, older larvae decrease their tail beats per bout but compensate by swimming at a faster overall pace (shorter resting periods between bouts). In vision-only conditions, older larvae compensate for fewer tail beats by increasing their tail curvature (swimming harder per beat).
Ecologically, this algorithm drives the fish away from the shorelines and into the center of streams where the flow is faster but the gradient is flat.Because steep gradients automatically form around solid objects in a current, this gradient-avoidance behavior inherently acts as a mechanism for obstacle avoidance.
References:
- Kulpa, M., Bak-Coleman, J., & Coombs, S. (2015). The lateral line is necessary for blind cavefish rheotaxis in non-uniform flow. The Journal of Experimental Biology.
- Olive, R., Wolf, S., Dubreuil, A., Bormuth, V., Debrégeas, G., & Candelier, R. (2016). Rheotaxis of Larval Zebrafish: Behavioral Study of a Multi-Sensory Process. Frontiers in Systems Neuroscience, 10:14. https://doi.org/10.3389/fnsys.2016.00014.
- Olszewski, J., Haehnel, M., Taguchi, M., & Liao, J. C. (2012). Zebrafish Larvae Exhibit Rheotaxis and Can Escape a Continuous Suction Source Using Their Lateral Line. PLoS ONE, 7(5), e36661. https://doi.org/10.1371/journal.pone.0036661.
- Oteiza, P., Odstrcil, I., Lauder, G., Portugues, R., & Engert, F. (2017). A novel mechanism for mechanosensory-based rheotaxis in larval zebrafish. Nature, 547, 445–448. https://doi.org/10.1038/nature23014.